
Interactions among Multiple Mutualists
Understanding the ecology, evolution, and molecular basis of mutualist diversity
In nature, most organisms interact simultaneously with multiple mutualistic species (e.g., plants with pollinators, microbial symbionts, and ants), however mutualism studies have traditionally focused on bipartite interactions between a host and a single partner (or type of partner). Bipartite studies will underestimate the importance of mutualism for ecological and evolutionary dynamics if complementarity occurs among functionally different partners, and overestimate it if different partners are in conflict. In a synthesis paper (Afkhami et al. Ecology 2014), we merged two important, but disparate, research approaches (network and consumer-resource models) to define the multiple mutualist effects – MMEs – that occur when a focal species has multiple partner mutualists. We have continued to think about what factors drive mutualist diversity (Batestone et al. Ecology 2018) and the importance of MMEs across levels from genes to communities in recent years (lab-wide conceptual project: Afkhami et al. Current Opinion in Plant Biology 2020).

Medicago truncatula with small nodules 2.5 weeks after inoculation with rhizobia.

Medicago plants at 2.5 weeks: Left is a plant grown with both rhizobia and mycorrhizal fungi and right is a plant grown with neither.
In our empirical research, we conduct ecological experiments investigating MMEs on performance of organisms and have several ongoing projects aimed at interactions between microbes, plants, and pollinators. We have also begun to integrate the ecological conceptual frameworks with genomic approaches to provide new insights into the functional genomic basis of how organism balance and interact with multiple mutualists. For example, we have used factorial experiments, genome-wide association mapping, and RNAseq with the tri-partite mutualism between Medicago truncatula, rhizobia, and arbuscular mycorrhizal fungi to (1) identify genes uniquely expressed in the presence of multiple mutualists and (2) to detect polymorphisms associated with plant fitness in response to multiple mutualists. Our first paper looking at MME identified pervasive genome-wide effects of multispecies microbial effects on gene expression, including genes whose expression responds non-additively to multiple mutualists (Afkhami & Stinchcombe Molecular Ecology 2016). To see a great summary article on our Molecular Ecology paper click here. To embrace both ecological and molecular complexity, we then expanded this work to incorporate gene-gene interaction asking how multispecies mutualisms impact coexpression network structure of their host plants. This project was led by University of Miami undergrad Sathvik Palakurty (Palakurty et al. Molecular Ecology 2018).
Most recently, we demonstrated that Multiple Mutualism Effects can also impact evolutionary trajectories of organisms by generating synergistic selection on plant traits and strengthening fitness alignment in the interaction between legumes, rhizobia and mycorrhizal fungi (Afkhami et al. Ecology Letters 2021).
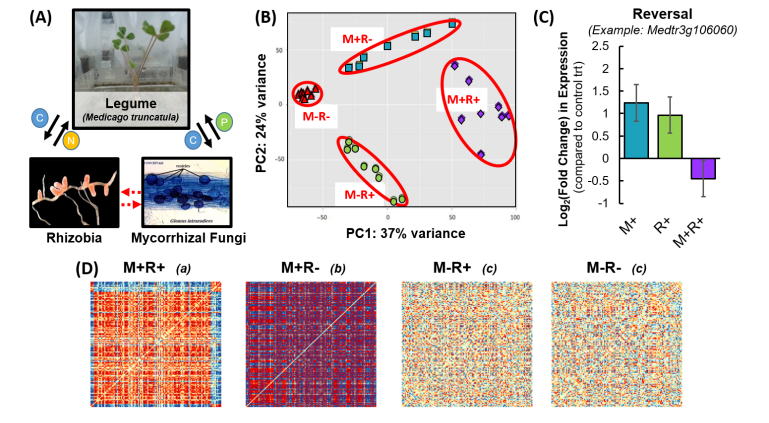
Experiments manipulating multispecies microbial mutualisms between plants, nitrogen-fixing bacteria, and mycorrhizal fungi (A) demonstrated significant changes in the genomewide expression (B) and gene expression networks (D) of host plants and identified candidate genes underlying plant regulation of multispecies mutualisms and synergistic plant growth (C). Subset of results from Stinchcombe & Afkhami 2016, Palakurty et al 2018, and Hernandez et al 2019. Please see papers for details.
Microbiomes and Plant-Microbe Interactions Under Stress
How does human-driven stress and disturbance impact microbes and interactions with plants?
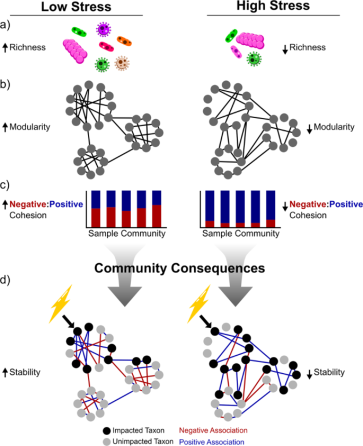
From Hernandez et al ISME 2021: The role for environmental stress in destabilization of microbial networks. Communities in high-stress environments (right) compared to communities in low stress environments (left) have less stable network properties. Communities under high stress are less species rich (a), less modular (b), and are less dominated by negative associations (lower negative:positive cohesion, c). The consequences for microbiomes (d) are that the reduction or loss of a taxon in response to an environmental disturbance (lightning bolt) can more easily propagate to the rest of the community (impacted taxa in black, unimpacted in gray). Positive associations (blue) are important pathways through which the effects on one taxon can cascade to impact other taxa in the community because: (1) the loss of a facilitator reduces the facilitated taxon’s fitness and/or (2) positively associated taxa are likely to be impacted by the same environmental factors (i.e., their niches strongly overlap).
In the Anthropocene, growing human populations intensify environmental stressors, threatening biodiversity and species interactions that underpin ecosystem function and productivity. Microbiomes can provide substantial benefits to the organisms that host them, ameliorating stresses and potentially increasing resilience. In many cases, however, it remains unclear how these microbial communities are affected by anthropogenic stresses, and what if any role they play in mitigating or exacerbating host responses to these stresses. Because of South Florida’s location and exploding human population, essentially all ecosystems in our region are imperiled and face anthropogenic stresses that stem largely from urbanization and climate change. Thus, we are very interested in informing how stress impacts the microbial communities, plant-microbial interactions, and the ecosystem services they provide, and we use a wide range of tools toward those goals (including experiments, microbiome sequencing, metatranscriptomics, etc).
For instance, graduate student Damian Hernandez recently led a paper showing that “Environmental stress destabilizes microbial networks” (Hernandez et al. ISME 2021). By assessing changes in diversity and structure of soil microbiomes along 40 replicate stress gradients in the Florida scrub ecosystem, he provided the first evidence that naturally-occurring microbiomes display network properties characteristic of unstable communities when under persistent stress. Further, graduate student Kasey Kiesewetter recently showed for the first time that habitat fragmentation — the breaking up of natural habitats with human-modified lands — is causing changes within the microbiome that in turn affects plant performance (Kiesewetter and Afkhami New Phytologist 2021). In addition, postdoc Suresh Subedi led a paper testing how the salinity stress caused by sea level rise impacts the interaction between mangroves and foliar endophyte microbiome, finding that salinity-experienced microbiomes are better able to confer salinity-tolerance to this important coastal tree. Other research led by postdoc Daniel Revillini investigated how fire stress not only impacts microbiome composition and interactions with native plants (Revillini et al. New Phytologist 2021), but also microbial community functional responses by using metatranscriptomics to identify shift in functional gene expression after fire (Revillini et al. in prep).

Picture from LILA where we do Everglades research. LILA a living laboratory in run by the water management district where restoration practices can be tested on a landscape scale.
We also work with local agencies to do research relevant to applied management and restoration efforts. For example, we work with the South Florida Water Management District on several Everglades projects investigating how hydrological management plans and restoration decision on tree islands impact plant-microbiome interactions important for restoring this unique and human impacted system (e.g. Almeida et al. Diversity 2021; Almeida et al. Restoration Ecology in review).
.
.
.
.
Mutualisms and Microbiomes Effects on Species Distributions
Do microbial landscapes structure plant distributions?

Figure 2 from Afkhami et al 2014 Ecology Letters: (A) Species distribution models demonstrated the geographic differentiation between fungal-associated (E+) and fungal-free (E-) Bromus laevipes. Habitat suitability for B. laevipes ranged from high (red) to low/unsuitable (blue). (B) E+ B. laevipes occupied a different climatic niche than E- populations, including uniquely occupying dry habitats (low values of PC3). Endophyte-associated changes to the climatic niche occurred along PC2-4, which explained ca. 45% of the variation in California’s climate. Filled circles = E+ populations, open circles = E- populations.
Understanding the factors underlying species distributions is fundamental to advancing ecological and evolutionary theory and to achieving conservation goals. Such an understanding can elucidate drivers of biodiversity, forecast species extinctions, and quantify invasion potential of introduced species. Recent work has produced new theory on the roles of biotic interactions in shaping species’ distributions, but these effects are rarely investigated over large geographic scales. Unlike antagonistic species interactions which contract a species’ range, positive species interactions have the unique potential to ameliorate biotic and abiotic stressors, and may thus expand species niches, resulting in larger species ranges.
In a one paper, we demonstrated that associating with a facultative mutualist can increase the range of a native species by thousands of square kilometers, or approximately 20% of the entire host range (Afkhami et al. Ecology Letters 2014). Combining (1) range-wide field observations from ~100 populations, (2) species distribution models, (3) 10 common garden experiments across the species range, and (4) greenhouse experiments, we showed that endophytic fungi can ameliorate drought stress and allow their grass host (Bromus laevipes) to grow in drier regions unoccupied by grasses that lack this partner. Range divergence between B. laevipes plants with and without endophytes was comparable to species-level divergence among congeners (other native Bromus), indicating that the mutualists’ effects were biologically significant. (To see this great summary article on our paper click here.)
Alterations to species niches and/or ranges caused by mutualisms have the potential to also play an important role in the evolution and diversification of species. Mutualist-generated niche differentiation could pave the way for speciation, if selection diverges across niches, leading to reduced gene flow. Collaborators and I are examined how association with mutualistic nitrogen-fixing bacteria affects diversification rates across ~20,000 species of legumes, the third most speciose family of plants (Afkhami et al. Ecology 2018). I am now working on how seed-borne fungal endophytes affect diversification in all of angiosperms as part of a recently funded NSF Dimensions of Biodiversity grant, collaborating with institutions here in the US and in China.
To scale microbe-mediated effects up to whole microbial communities (rather than individual microbes) and broader plant communities (rather than a single focal species), we have worked to develop a new system in the Florida Scrub at Archbold Biological Station. This system allows us to combine long-term research and monitoring of plant species at landscape scales (e.g., NSF LTREB data) with new microbiome experiments, microbial metagenomics/transcriptomics, and modeling.
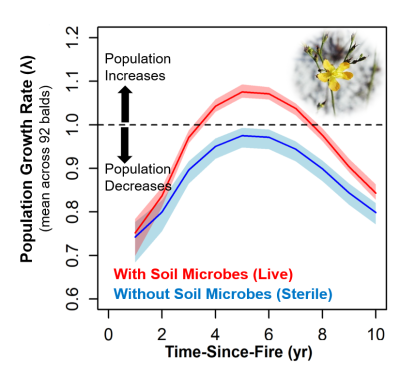
Population persistence of endangered plant Hypericum cumulicola depended on its soil microbiome across 92 sites. Integral Project Models parameterized with demographic data from experiments and the field showed that plant population growth rates only exceeded the threshold of λ=1 (i.e. the plant population was growing) when the appropriate soil microbiome was present. Population growth rates are shown projected along a fire history gradient (i.e. time since most recent fire). Figure from David et al 2019).
While studies have shown strong effects of microbiomes on individual-level fitness of plants, it remains unclear how these effects scale up to impact population and metapopulation dynamics. Understanding the consequences of microbiomes at these levels of biological organization is required for a mechanistic understanding of how species distributions are impacted by microbial associations. We received an NSF grant (“Microbial landscapes: Are microorganisms hidden drivers of species distributions?”) to examine how microbiomes effects on individual level plant fitness scale up to impact population, metapopulation, and ultimately landscape scale species distribution in the imperiled Florida Scrub ecosystem.
Work in this system by postdoctoral researcher Aaron David and post-baccalaureate student Khum Thapa Magar led to the development of the “Microbial Mitigation to Exacerbation Continuum”, which is a framework for understanding microbiome mediation of environmental stress and incorporates microbiomes into the Stress Gradient Hypothesis framework. We demonstrate that the interaction between soil microbiomes and environmental stress gradients is highly predictive of landscape scale distributions of threatened and endangered plant species (>80% of the variation in plant species occurrence) (David et al. Ecology 2018). In related work, we tested for the first time whether habitat specialization of the host plant affects adherence of microbiome-plant interactions to predictions of the stress gradient hypothesis (David et al. Ecology 2020). Further, a project combining integral projection models and manipulative experiments showed that the microbiome was required for population persistence of an endangered plant across the landscape and explained 47% of the variation in population growth rates among sites, substantially more than that attributable to abiotic variables (David et al. American Naturalist 2019).
Keystone Microbial Mutualists
Which hidden players structure plant and microbial communities?

This is the Florida Rosemary Scrub habitat which is highly imperiled (~15% of the historic distribution remaining). We are studying whether putative keystone microbes impact soil microbial community assembly after fire here.
Keystone species are important for understanding the maintenance of biodiversity and alternative stable states, as well as in guiding decisions on the conservation and restoration of ecosystems. While studies of keystone species have focused on highly visible macro-organisms, we have shown that microfungal endophytes can function as keystone mutualists influencing plant community structure. A 3-year field manipulation of endophyte in a native Californian grass showed that high endophyte abundance dramatically increased plant community diversity and altered community composition by suppressing a dominant invasive plant. The quantitative effects of endophytes on community diversity was on par with textbook examples of keystone species, such as starfish in the intertidal. Moreover, endophytes increased plant community diversity in 5 additional common gardens spanning >400km across the host grass range, demonstrating the keystone role of these fungi occurs on a range-wide scale(Afkhami & Strauss, Ecology, 2016).
Graduate Student Amanda Rawstern is now taking the next step to look at keystone microbes responsible for structuring the microbiome itself. Specifically, she is testing if putative “keystone microbes” identified from microbiome networks can actually structure microbiome community assembly in the Florida Scrub ecosystem at Archbold Biological Station.
Cascading effects of plant-microbial interactions
How do plant-microbial interactions impact other members of the community?
We are interested in accessing how plant-microbial interactions influence other members of the community, such as pollinators and herbivores. Some of our early work has shown how foliar fungal endophytes can deter natural enemies, reducing damage to plant hosts (e.g., Afkhami & Rudgers Environ. Entomology 2009), and genetic and chemical evidence has showed that these can endophytes produce a diversity of anti-herbivore alkaloids (Charlton, Craven, Afkhami et al. FEMS Microbiology Ecology 2014). We are now looking at how foliar endophyte diversity impacts plant investment in pollination and how habitat fragmentation influences the interaction between foliar endophyte communities, plant communities, and herbivore communities in the Pine Rocklands.

Pictures of the highly imperiled Pine Rocklands (2% of historic distribution remaining) and natural enemy damage on native plants.
Conflict in Positive Interactions
What are the sources and consequences of conflict in ‘beneficial’ interactions?
Vertically transmitted symbionts (i.e., passed parent to offspring) associate with some of the most ecologically dominant species on Earth, and their fixation has

Cheaters must prosper: Reconciling theoretical and empirical perspectives on cheating in mutualism. (Ecology Letters, 2015)
led to major evolutionary transitions (e.g., chloroplasts). Theory predicts that vertically-transmitted symbionts will be highly mutualistic because the fitness of the symbiont and host are coupled. However, partner conflict still exists in these intimate associations.
(1) Jenn Rudgers and I documented the first example of imperfect vertical transmission in the widespread symbiosis between plants and fungal endophytes (Afkhami & Rudgers American Naturalist 2008), which has since been identified as a common phenomenon and a central cause of variation in endophyte frequencies in nature (Rudgers, Afkhami et al. Ecology 2009).
(2) In a collaboration with Jenn Rudger’s lab, we documented subtle conflict due to the maternal inheritance of symbiosis, which biased host sex allocation toward maternal seed production at a cost to the host (Gorischek, Afkhami et al. American Naturalist 2013). To our knowledge this is the first example of a
symbiont altering sex allocation in plants.
(3) As part of an NCEAS working group, I have been collaborating on a novel
synthesis of the empirical and theoretical literature on cheating in mutualisms (Jones, Afkhami, et al. Ecology Letters 2015).
